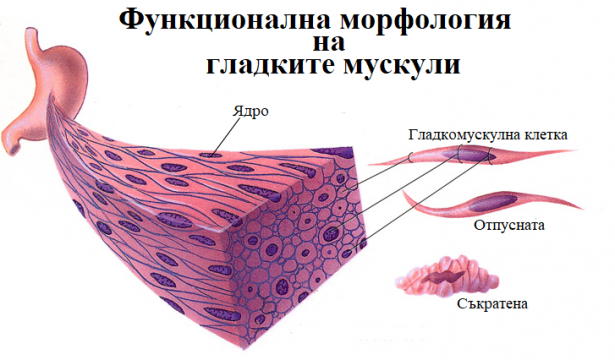
Функционална морфология на гладките мускули
Мускулите са активната част на опорно-двигателната система. Въз основа на функционалните и структурните си особености мускулите се делят на скeлетни мускули, гладки мускули и сърдечен мускул. Скелетните мускули и сърдечния мускул под микроскоп са напречонабраздени. Гладките мускули изграждат стените на вътрешните органи и не са напречнонабраздени. Приликите между тях и скелетните мускули са следните:
- и двата вида мускули генерират сила, която се стреми да намали дължината им;
- тази сила се поражда от миозиновите глави в резултат на тяхното взаимодействие с актина и хидролизата на АТФ;
- тези взаимодействия се извършват само когато концентрацията на Ca2+ в саркоплазмата се повиши многократно.
 Гладките мускули са изградени от малки едноядрени клетки, наречени миоцити. За разлика от скелетните те не съдържат миофибрили и затова не са напречнонабраздени. Миоцитите имат вретеновидна форма. В средната си част са най-дебели като постепенно изтъняват към краищата. Дължината на клетките варира в отделните тъкани. Най-късите се намират в кръвоносните съдове, а най-дългите в бременната матка. Гладките мускули не притежават сухожилия. Техните клетки са подредени на слоеве, които изграждат стените на кухите органи. Стените на някои вътрешни органи са изградени от повече от един слой гладка мускулна тъкан, като ориентацията на клетките в единия и в другия слой е различна. Така например стената на тънкото черво е изградена от два гладкомускулни слоя – вътрешен и външен. Влакната на вътрешния слой са разположени напречно (циркулярно), а на външния са разположени надлъжно. Миоцитите са покрити от базална мембрана. На места тя е прекъсната и съседните клетки се свързват с електрични синапси. В определени участъци на сарколемата гладкомускулните клетки са свързани механично.
Гладките мускули са изградени от малки едноядрени клетки, наречени миоцити. За разлика от скелетните те не съдържат миофибрили и затова не са напречнонабраздени. Миоцитите имат вретеновидна форма. В средната си част са най-дебели като постепенно изтъняват към краищата. Дължината на клетките варира в отделните тъкани. Най-късите се намират в кръвоносните съдове, а най-дългите в бременната матка. Гладките мускули не притежават сухожилия. Техните клетки са подредени на слоеве, които изграждат стените на кухите органи. Стените на някои вътрешни органи са изградени от повече от един слой гладка мускулна тъкан, като ориентацията на клетките в единия и в другия слой е различна. Така например стената на тънкото черво е изградена от два гладкомускулни слоя – вътрешен и външен. Влакната на вътрешния слой са разположени напречно (циркулярно), а на външния са разположени надлъжно. Миоцитите са покрити от базална мембрана. На места тя е прекъсната и съседните клетки се свързват с електрични синапси. В определени участъци на сарколемата гладкомускулните клетки са свързани механично.
Гладките мускули се характеризират с голямо разнообразие във функционално отношение. Тези, които изграждат матката, имат едни свойства, а тези, които регулират диаметъра на зеницата – съвсем други. Свойствата и особено регулацията на активността на гладките мускули са от голямо значение за медицинската практика. Редица заболявания, като артериална хипртония, бронхиална астма, дискинезия на жлъчните пътища, са свързани с нарушена функция на гладките мускули.
Сарколема
Клетъчната мембрана на гладкомускулните клетки се нарича сарколема. Тя не притежава напречни трансверзални каналчета (Т-каналчета) както напречнонабраздените мускули, но образува множество малки и плитки инвагинации, наречени кавеоли. Кавеолите са подредени в надлъжи ивици. Участъците, които не са снабдени с тях, образуват механични и електрични междуклетъчни свързвания. При някои гладки мускули електричните синапси са многобройни, като всеки миоцит е свързан чрез няколко електрични синапса със съседните клетки. Свързаните по този начин клетки функционират като едно цяло, те образуват един функционален синтициум. Гладките мускули от този тип се наричат висцерални, защото участват в изграждането на стените на вътрешните органи. Гладките мускули, които нямат или имат съвсем малко електрични синапси, се наричат мускули от дискретен тип. Такива са мускулите на окото, на семепроводите и mm. arrectores pilorum. При трети броят на електричните синапси не е постоянен, а се регулира хормонално. В матката по време на бременността почти не се срещат електрични синапси, но когато наближи раждането техният брой се увеличава, а след раждането отново намалява.
Сарколемата на миоцитите е снабдена още с йонни каналчета, йонни помпи и мембранни рецептори:

- йонни каналчета и помпи – сарколемата е снабдена с потенциалзависими Ca2+ каналчета. Те се наричат още дихидропиридинови рецептори, тъй като се блокират от вещества, спадащи към групата на дихидропиридините. Тези рецептори започват да се активират при мембранен потенциал около 30mV и остават отворени сравнително дълго време, поради което се наричат L-тип Ca2+. При някои гладки мускули разтягането на миоцитите води до повишаване на пропускливостта на сарколемата за калциеви йони. Това се дължи на наличието на механозависими Ca2+ каналчета. Сарколемата притежава още потенциал зависими закъсняващи К+ каналчета, но не притежава потенциал зависими Na+ каналчета. Освен до сега изброените сарколемата на гладките миоцити има лигандзависими Ca2+ каналчета и лигандзависими К+ каналчета. Активният транспорт през сарколемата се извършва през натриево-калиева помпа. Последните два транспортни механизма поддържат ниска стойност на калций в цитоплазмата.;
- мембранни рецептори и хормони – гладките мускули се инервират от вегетативната нервна система. При висцералните миоцити се откриват мембрани рецептори за медиаторите на вегетативната нервна система, които са пръснати почти по цялата сарколема. Един миоцит притежава няколко вида рецептори. За гладките мускули от дискретен тип е характерно, че рецепторите са концентрирани в областта на постсинаптичната мембрана. Сарколемата на гладкомускулните клетки притежава и рецептори за някои хормони (окситоцин, антидиуретичен хормон, ендотелин, ангиотензин II). При свързването на хормона с рецептора в по-голяма част от случаите се наблюдава засилване на съкращението на миоцитите, но може да се предизвика и тяхното отпускане.
Саркоплазмен ретикулум
 Саркоплазменият ретикулум е изключително важен клетъчен органел за осъществяване на мускулното съкращение. Той се състои от множество каналчета и везикули, които са пръснати из цялата саркоплазма. Тяхната гъстота е най-голяма в близост до участъците, където има множество кавеоли. Стената на саркоплазмения ретикулум е изградена от двоен липиден слой, в който са интергрирани множество протеини, които функционират или като Ca2+ помпа или като лигандзависими Ca2+ каналчета. Калциевите каналчета са два вида. Едните могат да се свързват с веществото рианодин и се наричат рианодинови рецептори, а другите се отварят когато вторият посредник инозилтрифосфат (ИТФ) се свърже с тях и се наричат ИТФ-зависим Ca2+ каналчета. Рианодиновите рецептори се отварят в резултат на повишаване на концентрацията на калциевите йони в саркоплазмата. След тяхното отваряне от саркоплазмения ретикулум излиза значително количество калций. Другият вид каналчета се отварят когато към тях се свърже ИТФ. Той се получава в резултат на активиран на фосфолипаза С. Редица медиатори и хормони предизвикват съкращение на гладките мускули чрез инозитолтрифосфат.
Саркоплазменият ретикулум е изключително важен клетъчен органел за осъществяване на мускулното съкращение. Той се състои от множество каналчета и везикули, които са пръснати из цялата саркоплазма. Тяхната гъстота е най-голяма в близост до участъците, където има множество кавеоли. Стената на саркоплазмения ретикулум е изградена от двоен липиден слой, в който са интергрирани множество протеини, които функционират или като Ca2+ помпа или като лигандзависими Ca2+ каналчета. Калциевите каналчета са два вида. Едните могат да се свързват с веществото рианодин и се наричат рианодинови рецептори, а другите се отварят когато вторият посредник инозилтрифосфат (ИТФ) се свърже с тях и се наричат ИТФ-зависим Ca2+ каналчета. Рианодиновите рецептори се отварят в резултат на повишаване на концентрацията на калциевите йони в саркоплазмата. След тяхното отваряне от саркоплазмения ретикулум излиза значително количество калций. Другият вид каналчета се отварят когато към тях се свърже ИТФ. Той се получава в резултат на активиран на фосфолипаза С. Редица медиатори и хормони предизвикват съкращение на гладките мускули чрез инозитолтрифосфат.
Цитоскелет
Цитоскелетът се състои от плътни телца и междинни филамети (нишки). Той поддържа формата на гладкомускулната клетка. За цитоскелета се залавя контрактилния апарат на миоцита. Междинните филаменти са изградени главно от белтъка десмин. Плътните телца са с яйцевидна форма. Те са белтъчни структури, които съдържат α-актинин. Този белтък може да се свързва с положителния край на актиновите нишки. Плътните телца с гладкомускулната клетка може да са пръснати в цитоплазмата (цитоплазмени плътни телца) или да са здраво прикрепени към сарколемата (плътни зони). Между плътните зони на отделните миоцити съществува здрава механична връзка. Плътните телца са свързани по между си посредством снопчета от междинни нишки.
Контрактитен апарат
Контрактилният апарат на гладките миоцити се състои от дебели и тънки нишки. Дебелите нишки съдържат миозин, а тънките актин. Количеството на миозина в гладките мускули е 4 пъти по-малко отколкото в скелетните. Смята се, че отношението на броя на дебелите към броя на тънките нишки е 1:15. Миофиламентите не са правилно подредени в саркомери. Основните характеристики на миофиламентите са:
- дебели (миозинови) нишки – те са изградени от две изоформи на миозин II. Той прилича на миозина на скелетните мускули, но се различава по аминокиселинния си състав. Всяка глава на миозина има по две леки вериги – една основна и една регулаторна. Активността на миозиновата АТФаза е многократно по-ниска. Фосфорилирането на регулаторната верига води до значително повишаване на активността и. Само след това активиране миозиновата глава може да се свърже с актина.
 Дебелите нишки са изградени от голям брой миозинови молекули. При напречен срез гладкомускулната дебела нишка има форма на правоъгълник, като миозиновите глави лежат само от двете срещуположни страни на квадрата, а другите две страни нямат глави. Опашките на миозиновите молекули са успоредни. Половината от главите са обърнати надясно, а другата половина – наляво. По този начин се получават две успоредни редици от миозинови глави. Плътната структура, която се образува от миозиновите опашки, се нарича стол на дебелата нишка. Десните глави могат да се извиват и дърпат актиновите нишки надолу, а левите – нагоре. Гладкомускулната дебела нишка има четири стени, като глави имат само две от стените. Всички миозинови молекули, чиито глави са разположени по едната стена на нишката, имат еднаква ориентация в пространството. Този начин на подреждане се означава като странична полярност.
Дебелите нишки са изградени от голям брой миозинови молекули. При напречен срез гладкомускулната дебела нишка има форма на правоъгълник, като миозиновите глави лежат само от двете срещуположни страни на квадрата, а другите две страни нямат глави. Опашките на миозиновите молекули са успоредни. Половината от главите са обърнати надясно, а другата половина – наляво. По този начин се получават две успоредни редици от миозинови глави. Плътната структура, която се образува от миозиновите опашки, се нарича стол на дебелата нишка. Десните глави могат да се извиват и дърпат актиновите нишки надолу, а левите – нагоре. Гладкомускулната дебела нишка има четири стени, като глави имат само две от стените. Всички миозинови молекули, чиито глави са разположени по едната стена на нишката, имат еднаква ориентация в пространството. Този начин на подреждане се означава като странична полярност.
Не всички миозинови молекули са включени в състава на дебелите нишки. Част от тях се намира в саркоплазмата под формата на миозинови мономери. В миоцитите постоянно протичат два противоположни процеса – полимеризация на миозиновите молекули, при която се получават повече и по-дълги миозинови нишки, и деполимеризация, при която нишките се разграждат до миозин-мономер. Ако мускулът бъде активиран преобладава полимеризацията. Така съкрашенията водят до увеличаване на броя и дължината на дебелите нишки.
Миозиновите нишки в гладките мускули са динамични структури. Те непрекъснато се разграждат и отново се изграждат. Тази динамика улеснява приспособяването на контрактилния апарат към дължината на миоцита.
- тънки (актинови) нишки – те са изградени от G-актин. Отделните молекули се свързват една с друга, като изграждат двойна спирала, която е основата на тънката нишка. Гладкомускулните актинови нишки са доста дълги. Тяхната дължина варира в широки граници. Подобно на миозиновите нишки и актиновите са динамични структури. В зависимост от условията броят и дължината им се променя. Тънките нишки на гладките мускули имат тропомиозин, но не притежават тропонин и небулин. Вместо тропонин те съдържат други два регулаторни белтъка – калдесмон и калпонин. Тропомиозинът е разположен надлъжно по повърхността на двойната актинова спирала, като покрива активните центрове на 7 актинови молекули. По време на съкращение той се премества в брадата между две актинови молекули. При гладките мускули неговото положение се регулира от съвместното действие на миозина и калдесмона. Миозиновите глави бутат тропомиозина навътре към браздата. Така миозинът се опитва сам да освободи активните центрове на актина, за да се върже с тях. Калдесмонът бута тропомиозина в обратна посока и не му позволява да се премести към браздата. Той е нишковиден белтък, който е надлъжно прикрепен към двойната актинова спирала. С единия си край той се свързва с актина, а с другия с миозина. Калдесмонът притежава залавни места за тропомиозина и за комплекса Ca2+-калмодулин. Той пречи на мускулното съкращение и понижава активността на миозиновата АТФаза. Другият регулаторен белтък е калпонин. Той има значително по-малки размери. Калпонинът е свързан с актина, но може да се свързва и с тропомиозина. Молекулата на този белтък притежава едно залавно място и за комплекса Ca2+-калмодулин. Калпонинът понижава активността на миозиновата АТФаза, като пречи по някакъв начин на активиращото действие на актина върху миозина.
Гладките мускули нямат саркомери. В тях има образувани съставени от актинови и миозинови нишки, които приличат в известна степен на саркомер. По тази причина някои ги наричат минисаркомери. Един минисаркомер е съставен от две съседни плътни телца и миофиламентите, разположени между тях. Плътните телца могат да се разглеждат като аналог на Z-дисковете при скелетните мускули. От тях излизат снопчета от актинови нишки, като всяко едно снопче е изградено от 20-25 актинови нишки с различна дължина. Между актиновите се намират 3-5 миозинови нишки. Миозиновите и актиновите нишки са полярни структури. Когато миозиновите глави се свържат с актина, те дърпат тънките нишки и дават възможност на минисаркомера да се скъси.
Гладките мускули са изградени от вретеновидни клетки, които нямат напречна набразденост. При тях липсват саркомерите, системата от Т-каналетата и регулаторният белтък тропонин. Въпреки разликите в структурата на гладките и на напречнонабраздените мускули, съкращението на гладките отново се осъществява чрез приплъзване на актиновите спрямо миозиновите нишки.


Продукти от Framar.bg

СПАЗМАЛГОН КОМПЛЕКС прах за перорален разтвор в саше * 10
ДРОТАВЕРИН инжекционен разтвор 20 мг / мл 2 мл * 10 ДАНСОН
Библиография
http://cdn.intechopen.com/pdfs/37881/InTech-Structure_and_function_of_smooth_muscle_with_special_reference_to_mast_cells.pdf
https://biologydictionary.net/smooth-muscle/
https://opentextbc.ca/anatomyandphysiology/chapter/10-8-smooth-muscle/
Л. Витанова, Р. Гърчев; „Физиология на човека”; издателство АРСО
доц. Янков; "Физиология"
Arthur C. Guyton, M.D., John E. Hall, Ph.D.; „Text book of Medical Physiology“
Коментари към Функционална морфология на гладките мускули