Повърхностни структури на бактериалните клетки
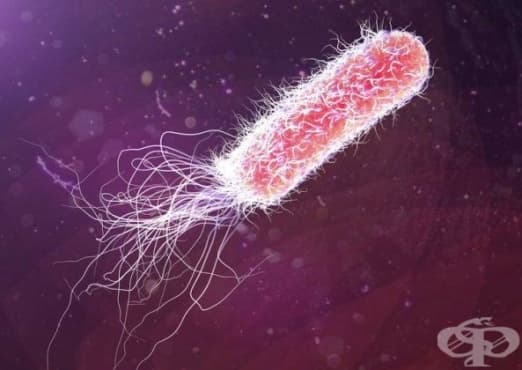
Бактериите са едни от най-успешните форми на живот, които обитават земното кълбо. Техният успех се дължи на простото им устройство и функция, както и на изключителната им способност за адаптиране към различни условия на заобикалящата ги среда. Тази им характеристика се подпомага от редица повърхностни структури, които ги защитават от вредни фактори на средата, позволяват придвижването и прикрепянето им към клетките на макроорганизмите, които населяват. Наред с капсулите и гликокаликса, други специфични повърхностни образувания на бактериалните клетки са пилите, ресните и аксиалните филаменти.
По повърхността на някои бактериални видове се откриват два вида клетъчни придатъци: ресни (флагели или камшичета), които представляват органи на движението, и пили (от лат. pilus - косъм), известни и като фимбрии. Камшичетата се срещат както при Грам-положителни, така и при Грам-отрицателни бактерии. За разлика от тях, пилите са характерни почти изключително за Грам-отрицателните видове, като изключение правят няколко грам-положителни микроорганизми (напр. Corynebacterium renale). Някои видове микроорганизми образуват на клетъчната си повърхност като ресни, така и пили.
Повърхностни структури на бактериалните клетки - пили
 Термините "пили" и "фимбрии", обикновено, се използват взаимозаменяемо, при описание на тънки, подобни на косъмчета придатъци по повърхността на множество Грам-отрицателни бактерии. Те представляват протеинови нишковидни образувания, кодирани от хромозомни и плазмидни гени, чиито белтъчни компоненти се наричат пилини. Те имат малки размери - диаметър от 2-8 nm и дължина 0,2-2 μm, и не служат за придвижване. Проксималният им край се вдава в цитоплазмената мембрана и всяка от тях е подредени около централен осев канал. Някои прокариоти притежават един тип пилин, докато други повече. Субединиците на отделните типове пили са съставени от различни протеини, което води и до разлики в антигенните им характеристики. По повърхността на една прокариотна клетка могат да се открият няколко типа фимбрии. Определени видове микроорганизми, като Shigella и E. coli, притежава до 200 пили по клетъчната си повърхност.
Термините "пили" и "фимбрии", обикновено, се използват взаимозаменяемо, при описание на тънки, подобни на косъмчета придатъци по повърхността на множество Грам-отрицателни бактерии. Те представляват протеинови нишковидни образувания, кодирани от хромозомни и плазмидни гени, чиито белтъчни компоненти се наричат пилини. Те имат малки размери - диаметър от 2-8 nm и дължина 0,2-2 μm, и не служат за придвижване. Проксималният им край се вдава в цитоплазмената мембрана и всяка от тях е подредени около централен осев канал. Някои прокариоти притежават един тип пилин, докато други повече. Субединиците на отделните типове пили са съставени от различни протеини, което води и до разлики в антигенните им характеристики. По повърхността на една прокариотна клетка могат да се открият няколко типа фимбрии. Определени видове микроорганизми, като Shigella и E. coli, притежава до 200 пили по клетъчната си повърхност.
В зависимост от функционалната си характеристика, пилите се делят на адхезивни и секс (полови) пили.
1. Адхезивните пили опосредстват прикрепването на бактериалните клетки към лигавиците на макроорганизма, което представлява първият етап на инфекциозния процес. Тази особеност ги определя като важни фактори на патогенността (болестотворност). Процесът на захващане на пилите към клетките на гостоприемника е специфичен и изисква наличието на комплементарен (съответстващ) рецептор по повърхността им. При отсъствие на съответния на адфезина рецептор, реализирането на инфекцията не е възможно. Адхезивните бактериални протеини могат да бъдат блокирани от специфични антитела (SIgA), като и от молекули, които съответства по химична структура на клетъчните рецептори. Фимбриите се свързват с рецептори, които по своето естество представляват олигозахариди.
Пилите на множество ентеробактерии придават на бактериалните клетки адхезивни свойства, които им позволяват да се прикрепят към различни епителни повърхности, червени кръвни клетки (причиняват хемаглутинация), както и към клетките на дрождите и гъбичките. Адхезивните (обикновени) пили тип 1 на представители от род Enterobacteriaceae притежават на върха си лектин (протеин), които се захваща специфично за D-маноза-съдържащи клетъчни рецептори при човека, животните, растенията и гъбите. Подобни тип 1 пили притежават уропатогенните колибактерии, клебсиела, ентеробактерии и други, които причиняват инфекции предимно на долните отдели на пикочните пътища (пикочен мехур, уретра), като и някои салмонели и холерния вибрион - за прикрепяне към ентероцитите на тънкото черво. Типа на пилите определя и специфичността на даден патоген към неговия хазяин.
Друг тип пили са т.нар P-пили на уропатогенни E. coli, които причинява инфекции на горните етажи на отделителната система. В клетките на бъбречните тъкани при човека се открива клетъчен рецептор, съдържа дигалактозен остатък, които е комплементарен на тези P-пили. Известен факт е, че единствено притежаващите адхезивни пили гонококи са патогенни. Така например Neisseria gonorrhea използват пили за хващане към сперматозоидите, чрез които се осъществява предаването им на друг човешки гостоприемник. Фимбриите на Грам-положителните пиогенни стрептококи са място на локализация на М-протеин, които представлява основният им повърхностен антиген. Именно на молекулната мимикрия на този антиген със структури на съединителната тъкан при човека, се дължи развитието на ревматизма. С фимбриите на тези стрептококи са асоциирани и липотейхоеви киселини, които опосредстват адфезията им към лигавичните епителни клетки. Адхезивните свойства на пилираните прокариотни клетки играят важна роля при бактериалната колонизация на епителните повърхности и следователно се определят и като фактори на колонизацията.
2. Секс пилите осъществяват конюгацията при Грам-негативните бактерии. Те позволяват прехвърлянето на генетичен материал между бактериалните клетки в процеса на конюгиране, поради което се наричат още и конюгиращи пили. Този процес играе съществена роля за предаване на лекарствената резистентност между бактериите. Секс пилите са от 1-4 четири на брой и са по-големи по размери в сравнени с обикновените пили. Може би най-добре проучен е пилусът на Escherichia coli, кодиран от т. нар. секс фактор F.
Секс пилусът обикновено е с диаметър от порядъка на 6-7 nm, а дължината му е 800-2000 nm. По време на конюгирането пилусът, излизащ от бактериалната клетка донор, захваща клетката на реципиента (получателя) и я привлича към себе си, при което се образува своеобразен свързващ мост. Последният осигурява директен контакт и образуване на пора, която позволява трансфера на ДНК от донора към реципиента. Обикновено, прехвърленият ДНК-фрагмент се състои от гени, необходими за създаване и трансфер на пили (често кодирани върху плазмид). Освен това, при този процес могат да бъдат трансферирани и други части на ДНК, което е причина за разпространение на генетични белези в бактериалната популация, като напр. антибиотична резистентност. Процесът на конюгация може да се осъществи и между бактерии от различни видове.
Други повърхностни фибрилерни структури, които са характери при Грам-положителните бактерии са липотейхоевите киселини и М-протеина на пиогенните стрептококи. Те представляват адхезини, чрез които бактериите се прикрепят към фибронектина - повърхностен гликопротеин на лигавичния епител. Грам-отрицателните бактерии не се свързват с фибронектина.
Повърхностни структури на бактериалните клетки - ресни
Ресните (флагели) или камшичетата представлява специфични извънклетъчни органели, чрез които някои видове пръчковидни и извити бактерии могат да се придвижват. Те са спирални полипептидни филаменти с дължина от порядъка на 5 до 12 μm и диаметър 12-30 nm. Под светлинен микроскоп могат да се наблюдават само след специфично оцветяване (по метод на Льофлер или Грей).

При синтезата на ресната отделните белтъчни субединици (флагелин) се сглобяват като формират цилиндрична структура с куха сърцевина. Отделните субединици се подреждат спирално и заграждат централен осев канал. Нарастването на ресната се извършва откъм дисталния й край, като отделните субединици преминават през централния канал и достигат нейния връх. Флагелумът се състои от три части:
1. Базално телце - разположено е в клетъчната обвивка. Състои се от стебло, което преминава през различен брой пръстени - при Грам-отрицателните са 4, а при Грам-положителните са 2. При Грам-отрицателните видове пръстените се намират в цитоплазмената мембрана, в пептидогликановия слой и във външната мембрана.
2. Кука - структура, разположена извън клетъчната обвивка, която се продължава от стеблото на базалното телце.
3. Филамент - дългата нишка, която започва след куката и се намира извън клетъчната повърхност. Флагелиновите субединици, които съставят филамента на ресната при бактериите са с изразени имуногенни свойства и са известни като H-антигени. В хода на инфекциозния процес срещу тях се изработват специфични антитела с важна защитна функция за макроорганизма. H-антигените се доказват чрез реакция аглутинация, която служи за серологично идентифициране на изолираните бактериални щамове в хода на микробиологичната диагностика. Бактериалните ресни денатурират и се разрушават при температура от 100 °C поради белтъчния си строеж.
В зависимост от броя и разположението на ресните могат да се различат няколко групи бактерии:
- монотрихи - бактерии, притежаващи една полярно разположена ресна (псевдомонади, вибриони);
- лофотрихи - притежават снопче от ресни, намиращо се в единия край на клетката (хеликобактер);
- амфитрихи - разполагат с една или две ресни от всеки и полюс на клетката (кампилобактер);
- перитрихи - ресните се разполагат по цялата клетъчна повърхност (ентеробактерии и бацили).
 Ресните служат като средство за придвижване на бактериалната клетка към източници на храна или други субстанции, както и за отдалечаването им от увреждащи ги вещества. Този процес е известен като химиотаксис. Движението на ресната се осигурява чрез въртене на базалното телце обратно на часовниковата стрелка, което предизвиква вихрова струя, осигуряваща подемна сила. Процесът е АТФ зависим и базалното тяло се задвижва от създаден протонен градиент при преминаване на протони през мембраната. Бактериите, образуващи ресни могат да се предвижват предимно в течни и полутечни среди, като изключение от правилото е Proteus, чиито представители често пълзят върху използваните твърди среди. Те формират т.нар. H-колонии, наподобяващи кондензирана пара върху стъкло. На този феномен се дължи и наименованието на H-антигените при реснистите бактерии (от немски Hauch - дъх). Обратно на това, O-антигените на Грам-отрицателните бактерии, които не формират ресни, са наречени така от немското "без дъх" - ohne Hauch.
Ресните служат като средство за придвижване на бактериалната клетка към източници на храна или други субстанции, както и за отдалечаването им от увреждащи ги вещества. Този процес е известен като химиотаксис. Движението на ресната се осигурява чрез въртене на базалното телце обратно на часовниковата стрелка, което предизвиква вихрова струя, осигуряваща подемна сила. Процесът е АТФ зависим и базалното тяло се задвижва от създаден протонен градиент при преминаване на протони през мембраната. Бактериите, образуващи ресни могат да се предвижват предимно в течни и полутечни среди, като изключение от правилото е Proteus, чиито представители често пълзят върху използваните твърди среди. Те формират т.нар. H-колонии, наподобяващи кондензирана пара върху стъкло. На този феномен се дължи и наименованието на H-антигените при реснистите бактерии (от немски Hauch - дъх). Обратно на това, O-антигените на Грам-отрицателните бактерии, които не формират ресни, са наречени така от немското "без дъх" - ohne Hauch.
Броят и разпределението на флагелите по бактериалната повърхност са характерни за даден вид и следователно са полезни при идентифицирането и класифицирането на бактериите. Например V. cholerae има единичен флагелум при единия полюс на клетката (т.е. монотрихи), докато Proteus vulgaris и Е. coli имат множество ресни, разпределени по цялата клетъчна повърхност (перитрихи). Важно е да се отбележи, че флагелите не са постоянен видов белег и при различни условия могат да липсват. Те могат да бъдат отстранени от клетъчната повърхност, без да се засяга жизнеспособността на клетката, което може да се осъществи чрез разбъркване на бактериалната култура със стъклени перли или в специален агар. След това клетката става временно неподвижна. С течение на времето се синтезира нова фланела и способността за движение на бактериите се възстановява. Инхибиторът на протеиновия синтез - хлорамфеникол, блокира регенерирането на ресните.
Повърхностни структури на бактериалните клетки - аксиларни филаменти
Движенията на спирохетите - борелии, трепонеми, лептоспири, се създават от т.нар. аксиларни или осеви филаменти, които са съставени от белтъци, подобни на флагелин. Осевите филаменти се намират в периплазменото пространство на клетъчната обвивка, между цитоплазмената и външната мембрана. Те представляват паралелно вървящи и спирално навити нишки, които при различните видове варират на брой от 2-15 и повече. Филаментите завършват в краищата на клетките с т.нар. инсерционен апарат, състоящ се от кука и няколко инсерционни диска, които се поместват в клетъчната стена и плазмалема. Спирохетите се придвижват в течна и вискозна среда чрез три основни движение: ротации около надлъжната ос, сгъване и разгъване и придвижване напред-назад.
Библиография
https://www.ncbi.nlm.nih.gov/books/NBK8477/#A284
http://library.open.oregonstate.edu/microbiology/chapter/bacteria-surface-structures/
https://en.wikipedia.org/wiki/Bacterial_cell_structure#Extracellular_(external)_structures
Микробиология - Медицинско издателство "АРСО" София 2000
http://www.biologydiscussion.com/bacteria/bacterial-cells/bacterial-cell-structure-and-components-microbiology/54746
Коментари към Повърхностни структури на бактериалните клетки