Екстрацелуларен матрикс
В биологията, екстрацелуларния матрикс е колекция от извънклетъчни молекули, секретирани от поддържащи клетки, която осигурява структурна и биохимична подкрепа на заобикалящите клетки. Тъй като многоклетъчността еволюира независимо в различни многоклетъчни линии, съставът на екстрацелуларен матрикс варира между многоклетъчните структури.
Екстрацелуларният матрикс е не-клетъчна триизмерна макромолекулна мрежа, съставена от колагени, протеогликани/гликозаминогликани, еластин, фибронектин, ламинини и няколко други гликопротеини. Матричните компоненти се свързват помежду си, както и рецепторите за клетъчна адхезия, които образуват сложна мрежа, в която клетките се намират във всички тъкани и органи. Клетъчните повърхностни рецептори трансдуцират сигнали в клетките от екстрацелуларен матрикс, които регулират различни клетъчни функции, като преживяемост, растеж, миграция и диференциация и са жизненоважни за поддържане на нормална хомеостаза. Екстрацелуларният матрикс е силно динамична структурна мрежа, която непрекъснато претърпява ремоделиране, медиирано от няколко ензими, разграждащи матрицата по време на нормални и патологични състояния. Дерегулацията на състава и структурата му е свързана с развитието и прогресията на няколко патологични състояния.
Компонентите на екстрацелуларният матрикс се произвеждат вътреклетъчно от резистентни клетки и секретират в него чрез екзоцитоза. Веднъж секретирани, те се агрегират със съществуващата матрица. Екстрацелуларният матрикс се състои от свързваща мрежа от фиброзни протеини и гликозаминогликани.
Глюкозаминогликаните са въглехидратни полимери и най-вече се прикрепват към протеините на извънклетъчния матрикс, за да образуват протеогликани. Протегликаните имат нетен отрицателен заряд, който привлича положително заредени натриеви йони, които привличат молекули на водата чрез осмоза, като поддържат есктрацелуларен матрикс и живите клетки хидратирани. Протегликаните също могат да помогнат за улавяне и съхранение на растежните фактори в него.
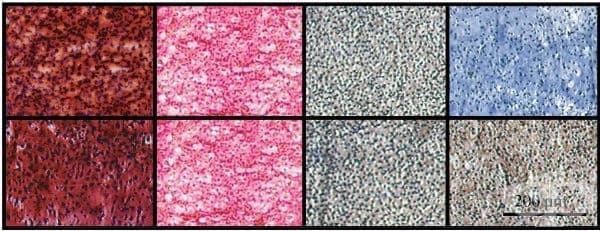
Гликозаминогликаните се идентифицират в тъканните срезове чрез различни хистохимични техники, включително оцветяване с алциново синьо и неговите аналози, като например куплолиново синьо и купромероново синьо или с високи и ниски методи на желязо диамин. Промяната в оцветяването води до особено объркване в случая на алцианово синьо, където се използват не само няколко различни марки алцианово синьо, но и няколко различни оцветяващи протокола. Ако резултатите от тези техники се сравняват, те често не съвпадат (фигура 1).
Еластините, за разлика от колагена, дават еластичност на тъканите, което им позволява да се разтягат при нужда и след това да се върнат в първоначалното си състояние. Това е полезно в кръвоносните съдове, белите дробове, в кожата и тези тъкани съдържат големи количества еластини. Еластините се синтезират от фибробласти и гладкомускулни клетки. Еластините са силно неразтворими и тропоеластините се секретират вътре в молекулата на протеин на чаперон, която освобождава прекурсорната молекула при контакт с влакно от зрял еластин. Тропоеластините след това се деаминират, за да се включат в еластиновата нишка.
Оцветяването на Verhoeff, известно също като еластично петно на Verhoeff или петно на Verhoeff-Van Gieson, е протокол за оцветяване, използван в хистологията, разработен от американския офталмолог и патолог Фредерик Херман Верхоф (1874-1968) . Формулата се използва за демонстриране на нормални или патологични еластични влакна.
Оцветяването на Verhoeff представлява голямо разнообразие от катионни, анионни и нейонни връзки с еластин, основната съставка на тъканта от еластични влакна. Еластинът има силен афинитет към желе-хематоксилиновия комплекс, образуван от реагентите в петна и следователно ще запази багрилото по-дълго от другите тъканни елементи. Това позволява еластинът да остане оцветен, докато останалите тъканни елементи са обезцветени (фигура 2). Натриев тиосулфат се използва за отстраняване на излишния йод и за контраст (най-често петното на Ван Гизон) се използва за контраст на основното петно. Еластичните влакна и клетъчните ядра са оцветени в черно, колагенните влакна са оцветени в червено, а други тъканни елементи, включително цитоплазмата, са оцветени в жълто.
Той заема най-големия дял от междуклетъчното вещество. Колагенът представлява група от близки, но генетично различни протеини, които имат общи свойства. Колагеновата молекула е формирана от три полипептидни вериги. Те са изградени от глицин, пролин и лизин, които образуват тройна спираловидна верига - алфа-верига. Някои видове колаген съдържат три еднакви a-вериги, например колаген тип III, а други като колаген тип I имат две еднакви и една различна вериги. Всяка полипептидна верига съдържа около хиляда аминокиселинни остатъка и има молекулно тегло около 35 000 D.
Полипептидните вериги на колагена се синтезират в рибозомите на фибробластите или подобни клетки в сухожилията, костите и хрущялите. След това навлизат в просвета на ендоплазматичния ретикулум под формата на про-алфа-вериги. Това не се отнася за колагена на базалните мембрани, който се синтезира от епителни и ендотелни клетки. В процеса на формиране на колагеновата верига участват голямо количество ензими като хидроксилази, пептидази и други доказано е, че при хипоксия, недостиг на желязо и аскорбинова киселина е невъзможно образуването на предшественика на колагена - проколаген.
Сириус червено е хистологично петно, използвано за маркиране на общия колаген, както и за различаване между различните видове колаген за оценка на разпределението на колаген в тъканите. Групата на сулфоновата киселина на Сириус червено реагира с основни аминогрупи на лизин и хидроксилизин и гуанидинова група на аргинин. По този начин, като анионно багрило, той се свързва с всички различни видове изоформи на колагена. В светло поле колагенът се появява като връзки от розово до червени влакна, които се разстройват в патологични условия. Същите по-големи колагенни влакна при поляризирана светлина изглеждат ярко жълто до оранжево, а по-тънките, включително ретикуларни влакна, изглеждат зелени (фигура 3). Това двойно пречупване, при което инцидентната светлина се разделя чрез поляризация на две различни пътища, е много специфична за колагена. Количеството на поляризираната светлина, абсорбирана от сириус-червеното багрило, зависи стриктно от ориентацията на колагенните снопове, позволяващи да се разграничат различните видове колаген.
Библиография
Color atlas of pathology, Section Cellular pathology
https://en.wikipedia.org/wiki/Extracellular_matrix
https://www.sciencedirect.com/science/article/pii/S0169409X15002574
https://www.ncbi.nlm.nih.gov/pubmed/7511937
https://en.wikipedia.org/wiki/Verhoeff%27s_stain
https://bio-protocol.org/e2592
Коментари към Екстрацелуларен матрикс